【实验研究】TGF-β/WNT5a/JNK信号通路对人晶状体上皮细胞上皮-间充质转化的促进作用
【摘要】目的 研究转化生长因子β/无翅蛋白5a/c-Jun N-氨基末端激酶(TGF-β/WNT5a/JNK)信号通路对晶状体上皮细胞(LECs)纤维化的影响。
方法 将人LECs细胞系SRA01/04分为对照组、TGF-β组和WNT5a组,其中对照组细胞常规培养,TGF-β组和WNT5a组分别采用TGF-β 1和WNT5a处理24 h。采用Western blot法检测3个组细胞WNT5a、JNK、磷酸化JNK(p-JNK)蛋白的相对表达量。另将细胞系分为对照组、TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组,其中对照组细胞常规培养,TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组分别采用TGF-β 1处理24 h、TGF-β 1处理24 h+JNK抑制剂SP600125处理2 h、WNT5a处理24 h+SP600125处理2 h。采用Western blot法检测4个组细胞WNT5a、JNK、p-JNK、Ⅰ型胶原蛋白(Col-Ⅰ)、纤维连接蛋白(FN)和α-平滑肌肌动蛋白(α-SMA)的相对表达量;采用免疫荧光染色法检测各组细胞α-SMA的表达分布;采用Transwell小室实验检测各组细胞相对迁移数;采用Col-Ⅰ凝胶收缩实验检测各组细胞培养8、16、24和48 h时Col-Ⅰ凝胶面积比率。
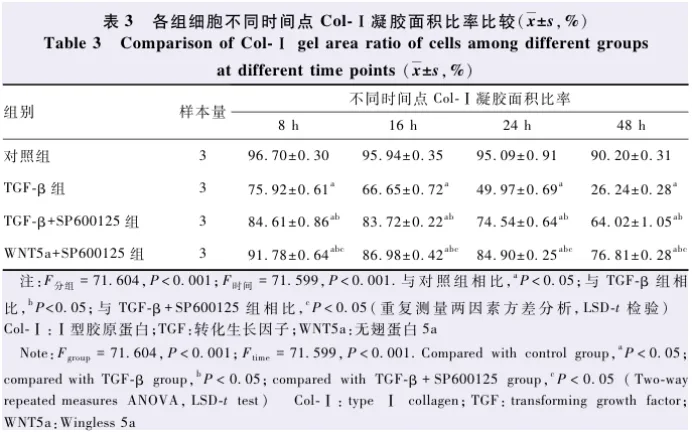
结果 Western blot结果显示,TGF-β组和WNT5a组细胞WNT5a、JNK、p-JNK蛋白相对表达量均明显高于对照组,差异均有统计学意义(均 P<0.05)。TGF-β组、TGF-β+SP600125组、WNT5a+SP600125组细胞Col-Ⅰ、FN、α-SMA的蛋白相对表达量明显高于对照组,TGF-β+SP600125组和WNT5a+SP600125组细胞各蛋白相对表达量明显低于TGF-β组,差异均有统计学意义(均 P<0.05)。免疫荧光染色结果显示,TGF-β组SRA01/04细胞由柱状的上皮细胞转分化为纺锤状的肌成纤维细胞,TGF-β+SP600125组和WNT5a+SP600125组细胞仍以柱状上皮细胞形态居多。TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞内α-SMA相对荧光染色强度明显高于对照组,TGF-β+SP600125组和WNT5a+SP600125组α-SMA相对免疫荧光强度明显低于TGF-β组,差异均有统计学意义(均 P<0.05)。Transwell小室迁移实验结果显示,TGF-β组细胞相对迁移数明显高于对照组,TGF-β+SP600125组和WNT5a+SP600125组细胞相对迁移数明显低于TGF-β组,差异均有统计学意义(均 P<0.05)。Col-Ⅰ胶原凝胶收缩实验结果显示,随着培养时间的延长,各组细胞Col-Ⅰ凝胶面积明显减小。各组不同时间点Col-Ⅰ凝胶面积比率总体比较,差异均有统计学意义( F分组=71.604, P<0.001; F时间=71.599, P<0.001);培养48 h时,TGF-β组、TGF-β+SP600125和WNT5a+SP600125组细胞Col-Ⅰ凝胶面积比率分别为(26.24±0.28)%、(64.02±1.05)%、(76.81±0.28)%,明显低于对照组的(90.20±0.31)%,差异均有统计学意义(均 P<0.05)。
结论 WNT5a/JNK信号通路作为TGF-β信号通路下游作用靶点,促进了LECs上皮-间充质转化及细胞外基质沉积,提高了细胞收缩力。
关键词:晶状体上皮细胞;c-Jun N-氨基末端激酶;转化生长因子-β;无翅蛋白5a;上皮-间充质转化;纤维化
基金项目:内蒙古自治区自然科学基金(2019MS08116);内蒙古医科大学面上项目(YKD2022MS021)
DOI: 10.3760/cma.j.cn115989-20230425-00147
白内障是常见的全球性致盲眼病之一,预计到2050年我国45~89岁人群的白内障患病率将占白内障患病总人群的33.34%[1]。手术是白内障的主要治疗手段。后发性白内障(posterior capsular opacification,PCO)是白内障术后常见的远期并发症,随着手术技术和人工晶状体的改良,PCO的发病率已有所下降,但术后2个月~5年成人和儿童PCO的总发病率分别高达50%和100%[2]。PCO的形成与囊膜下残留晶状体上皮细胞(lens epithelial cells,LECs)的增殖、迁移及上皮-间充质转化(epithelial-mesenchymal transition,EMT)密切相关。WNT信号通路是促进脊椎动物胚胎正常发育和维持成体组织内稳态的重要途径,WNT蛋白通过配体-受体结合,激活受体胞内结构域,将胞外信号转换为胞内反应。根据是否依赖β-连环蛋白,WNT信号通路分为经典和非经典信号通路,后者包括WNT/c-Jun N末端激酶(c-Jun N-amino terminal kinase,JNK)、WNT/Ca 2+通路[3]。大量体内外研究已证实,经典WNT信号异常激活与PCO直接相关,而非经典WNT信号在PCO中的作用机制却鲜有研究。WNT/JNK通过非经典WNT蛋白与RAR相关孤儿受体2、Vangl蛋白2和卷曲蛋白7(frizzled 7,FZD7)复合物结合,激活蓬乱蛋白介导的JNK、Rho/Rac GTP酶、Rho激酶信号,促进转录因子激活蛋白1和转录激活因子2的表达,从而调节细胞生物学行为[3,4]。在转化生长因子β(transforming growth factor-β,TGF-β)诱导的LECs EMT和白内障发生过程中,非经典WNT信号配体无翅蛋白5a(Wingless 5a, WNT5a)、受体FZD2等有所上调[5]。但WNT/JNK信号是否在LECs纤维化中起作用尚不清楚。本研究拟探索WNT/JNK信号通路对LECs纤维化的影响,以期为PCO的药物防治提供新的靶点。
1.1 材料
1.1.1 细胞来源
人LECs细胞系SRA01/04购自中国医学院肿瘤细胞库。
1.1.2 主要试剂及仪器
DMEM培养基、胎牛血清(fetal bovine serum,FBS)、青霉素-链霉素溶液(美国Gibco公司);3%牛血清白蛋白(bovine albumin,BSA)、荧光素偶联山羊抗小鼠IgG(A32723)、Western blot检测盒(美国Thermo Fisher公司);重组人TGF-β 1蛋白(P01137)、重组人WNT5a蛋白(P22725)、小鼠抗α-平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)单克隆抗体(MAB1420)、兔抗JNK多克隆抗体(AF1205)(美国D&R Systerm公司);兔抗WNT5a多克隆抗体(ab153876)、兔抗纤维连接蛋白(fibronectin,FN)单克隆抗体(ab199056)、JNK抑制剂SP600125、小鼠抗甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)单克隆抗体(ab181602)、兔抗WNT5a多克隆抗体(ab153876)、辣根过氧化物酶(horseradish peroxidase,HRP)标记山羊抗兔IgG(ab97051)、HRP兔抗小鼠IgG(ab6728)(美国Abcam公司);兔抗磷酸化JNK(p-JNK)多克隆抗体(9251)、兔抗Ⅰ型胶原蛋白(type Ⅰ collagen,Col-Ⅰ)多克隆抗体(42818)(美国Cell Signaling Technology公司);BCA蛋白试剂盒(美国Pierce公司);电化学发光液(美国Millipore公司);Ⅰ型鼠尾胶原(美国Corning公司);细胞收缩检测试剂盒(美国Cell Biolabs公司)。激光扫描共聚焦显微镜(德国Lecica公司);BS-300型全自动生化分析仪(深圳迈瑞生物医疗电子股份有限公司);化学发光检测系统(美国Tanon公司);DYY-11型多用电泳仪(北京六一仪器厂);MCO-175型CO2培养箱(日本三洋集团);光学倒置显微镜(IMF413,日本Olympus公司)。
1.2 方法
1.2.1 细胞体外培养
SRA01/04细胞在含10% FBS、100 U/ml(商品单位)青霉素和100 mg/ml链霉素的DMEM培养基中,于37 ℃、5% CO 2的条件下培养,每天换液。当细胞融合达80%时,加入1 ml胰蛋白酶消化液于37 ℃培养箱内消化2 min,按1∶4比例传代培养。取第5~7代细胞进行后续实验。
1.2.2 实验分组
将细胞接种至6孔板中,分为TGF-β组、WNT5a组和对照组3个组,分别采用2 ng/ml TGF-β1、5 ng/ml WNT5a和常规培养基培养24 h。重复接种新培养的细胞至6孔板中,分为4个组,其中TGF-β组细胞采用2 ng/ml TGF-β1诱导24 h;TGF-β+SP600125组细胞采用2 ng/ml TGF-β1诱导24 h后加入10 μmol/L JNK抑制剂SP600125继续处理2 h;WNT5a+SP600125组细胞采用5 ng/ml WNT5a诱导24 h后加入10 μmol/L SP600125继续处理2 h;对照组细胞常规培养24 h。
1.2.3 Western blot法检测各组细胞中WNT5a、JNK、p-JNK、Col-Ⅰ、FN及α-SMA蛋白表达量
取各组细胞,预冷PBS洗涤2次,每孔加入100 μl预冷SDS裂解液,4 ℃下12 000× g离心15 min,取上清获得细胞总蛋白,按照BCA蛋白定量试剂盒说明测定蛋白总浓度。选取35~50 μg蛋白上样到新配制的SDS-PAGE凝胶进行电泳,并转印至PVDF膜。室温下将PVDF膜置于5%脱脂奶粉中摇动封闭1 h,TBS洗涤3次;加入WNT5a、JNK、p-JNK、α-SMA、FN、Col-Ⅰ和GAPDH抗体(均1∶1 000稀释)于4 ℃下孵育过夜;TBS洗膜后与相应的HRP标记二抗(均1∶5 000稀释)室温振荡孵育2 h;ECL法显影,于暗室曝光并采用Tanon 5200化学发光成像仪进行拍照,采用ImageJ软件进行灰度分析,以GAPDH为内参照,计算各目的蛋白的相对表达量。实验独立重复3次。
1.2.4 免疫荧光染色法检测各组细胞中α-SMA表达分布
取各组细胞,以1×104/ml的密度接种于含玻片的24孔培养板中,置于培养箱中培养1~2 d,取出细胞爬片,用预热的pH 7.4 PBS洗涤2次,加入4%多聚甲醛固定15 min,PBS洗涤3次;用50 mmol/L NH 4Cl中和游离的多聚甲醛10 min,PBS洗涤3次,0.2% TritonX-100室温透膜5 min,PBS洗涤3次;加入3% BSA室温下封闭30 min;加入2 μg/ml α-SMA抗体(1∶1 000)4 ℃孵育过夜,PBS振荡洗涤3次;加入荧光素偶联二抗(1∶5 000)室温下孵育2 h,PBS洗涤3次;加入DAPI避光孵育5 min,PBS洗涤3次,用含抗荧光淬灭剂的封片液封片。采用激光扫描共聚焦显微镜观察并拍照,采用ImageJ软件分析各组细胞α-SMA相对荧光强度。实验独立重复3次。
1.2.5 Transwell小室实验检测各组细胞迁移情况
将8 μm孔径Transwell小室置于24孔板,上层加入0.05% BSA培养基,下层加入20% FBS培养基。分别取各组对数生长期的细胞,弃去培养基,PBS洗涤3次,加入胰蛋白酶消化至部分细胞呈球状,加入培养基轻轻振荡终止消化,吸出剩余培养基,PBS洗涤后用无血清培养基重悬,调整细胞密度为5×105个/ml。取200 μl重悬细胞加入上室聚碳酸酯膜上,37 ℃、5% CO2条件下孵育48 h。取出小室,棉签轻拭聚碳酸酯膜表面的细胞,4%多聚甲醛固定30 min,0.1%结晶紫染色20 min,PBS洗涤细胞3次,脱色,风干后在10倍视野显微镜下任意选取5个视野观察并计数。实验独立重复3次。
1.2.6 Col-Ⅰ凝胶收缩实验检测各组细胞胶原凝胶收缩情况
取1%BSA覆盖24孔板以孵化胶原凝胶,于37 ℃下培育1 h,避免凝胶与培养皿接触。将Ⅰ型鼠尾胶原配制成2 mg/ml中和胶原混合液。分别将4个组细胞用胰蛋白酶消化后用无血清DMEM/F12培养液重悬细胞并计数,将细胞悬液接种于Col-Ⅰ溶液中,调整细胞密度为2.5×105个/ml,每孔0.5 ml加入24孔板,37 ℃下静置1 h;37 ℃、5% CO2温箱中培育,分别于8、16、24和48 h监测胶原凝胶收缩情况。采用ImageJ软件分析胶原凝胶面积,计算各时间点胶原凝胶面积比率。各时间点胶原凝胶收缩面积比率=各时间点凝胶面积/初始凝胶面积。实验独立重复3次。
1.3 统计学方法
采用SPSS 26.0统计学软件进行统计分析。计量数据资料经Shapiro-Wilk检验证实呈正态分布,以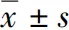 表示,组间经Levene检验证实方差齐。各组细胞WNT5a、JNK、p-JNK、Col-Ⅰ、FN、α-SMA蛋白相对表达量、相对细胞迁移数总体比较采用单因素方差分析,组间多重比较采用LSD- t检验。各组细胞不同时间点胶原凝胶收缩面积比率比较采用重复测量两因素方差分析,组间多重比较采用LSD- t检验。采用双侧检验, P<0.05为差异有统计学意义。
表示,组间经Levene检验证实方差齐。各组细胞WNT5a、JNK、p-JNK、Col-Ⅰ、FN、α-SMA蛋白相对表达量、相对细胞迁移数总体比较采用单因素方差分析,组间多重比较采用LSD- t检验。各组细胞不同时间点胶原凝胶收缩面积比率比较采用重复测量两因素方差分析,组间多重比较采用LSD- t检验。采用双侧检验, P<0.05为差异有统计学意义。
2.1 TGF-β组、WNT5a组和对照组细胞WNT5a、JNK、p-JNK蛋白表达情况比较
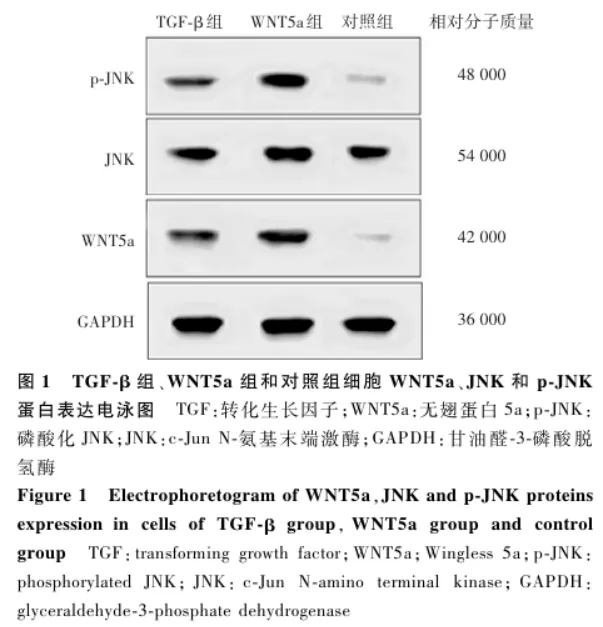
Western blot结果显示,TGF-β组和WNT5a组细胞WNT5a和p-JNK蛋白条带强度明显强于对照组( 图1 )。3个组细胞WNT5a、JNK和p-JNK蛋白相对表达量总体比较差异均有统计学意义( F=1 417.027、1 855.444、817.986,均 P<0.05),其中TGF-β组和WNT5a组细胞WNT5a、JNK和p-JNK蛋白相对表达量均高于对照组,WNT5a组各蛋白相对表达量均高于TGF-β组,差异均有统计学意义(均 P<0.05)( 表1 )。
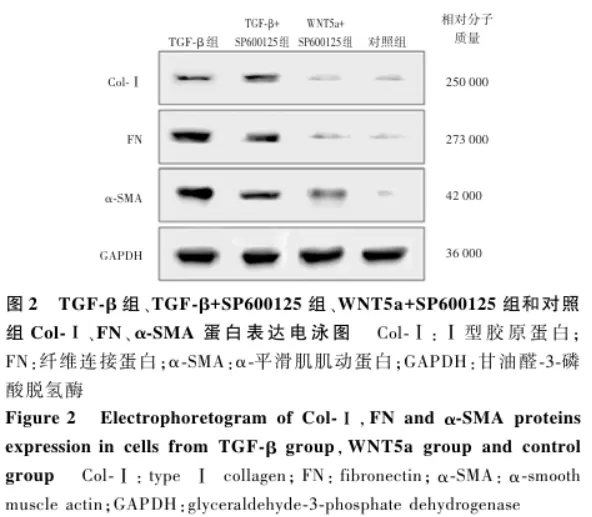
2.2 各组细胞Col-Ⅰ、FN和α-SMA蛋白表达情况比较
Western blot结果显示,TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞Col-Ⅰ、FN和α-SMA蛋白条带灰度强于对照组,TGF-β+SP600125组和WNT5a+SP600125组细胞3种蛋白条带灰度明显弱于TGF-β组( 图2 )。4个组Col-Ⅰ、FN和α-SMA蛋白相对表达量总体比较差异均有统计学意义( F=149.474、618.478、185.882,均 P<0.001),其中TGF-β组细胞Col-Ⅰ、FN和α-SMA蛋白相对表达量均高于TGF-β+SP600125组、WNT5a+SP600125组和对照组,TGF-β+SP600125组和WNT5a+SP600125组各蛋白相对表达量均高于对照组,TGF-β+SP600125组各蛋白相对表达量均高于WNT5a+SP600125组,差异均有统计学意义(均 P<0.05)( 表2 )。
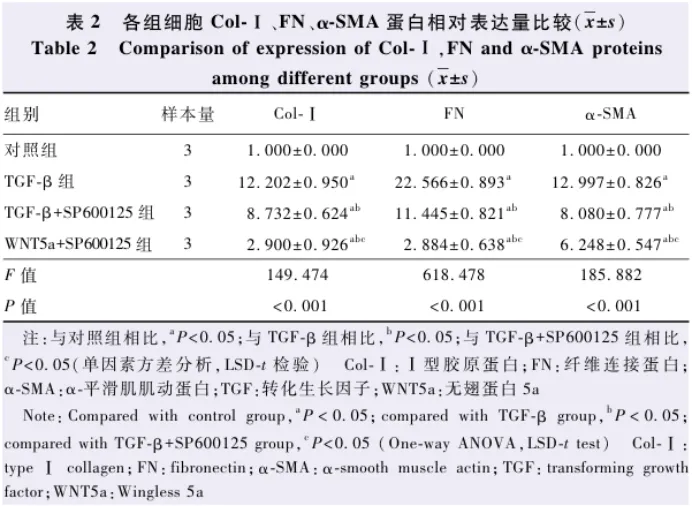
2.3 各组细胞α-SMA蛋白表达分布比较
与对照组相比,TGF-β组细胞由柱状的上皮细胞变为梭形的间质细胞。TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞α-SMA广泛分布在细胞核周边及细胞质中。TGF-β组、TGF-β+SP600125组、WNT5a+SP600125组和对照组α-SMA相对免疫荧光强度分别为53.627±3.067、42.083±1.744、22.743±1.635、0.000±0.000,总体比较差异有统计学意义( F=438.209, P<0.001),其中TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞α-SMA相对免疫荧光强度均明显高于对照组,TGF-β组细胞α-SMA相对免疫荧光强度明显高于TGF-β+SP600125组和WNT5a+SP600125组,TGF-β+SP600125组细胞α-SMA相对免疫荧光强度明显高于WNT5a+SP600125组,差异均有统计学意义(均 P<0.05)( 图3 )。
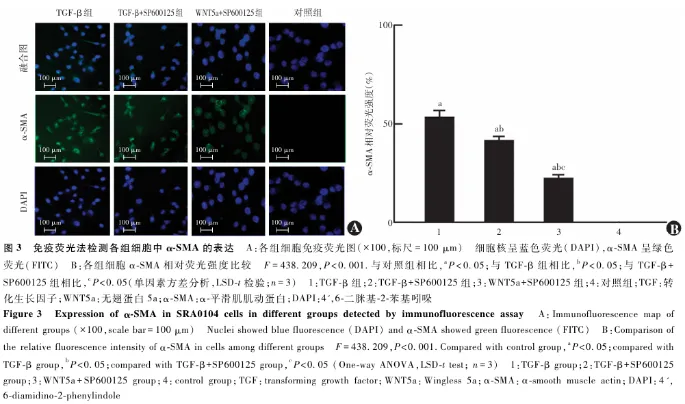
2.4 各组细胞迁移情况比较
Transwell小室迁移实验结果显示,TGF-β组、TGF-β+SP600125组、WNT5a+SP600125组和对照组细胞相对迁移数分别为4.86±0.14、2.86±0.14、2.29±0.14、1.00±0.14,总体比较差异有统计学意义( F=378.750, P<0.001),其中TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞相对迁移数均明显高于对照组,TGF-β组细胞相对迁移数明显高于TGF-β+SP600125组和WNT5a+SP600125组,TGF-β+SP600125组细胞相对迁移数高于WNT5a+SP600125组,差异均有统计学意义(均 P<0.05)( 图4 )。
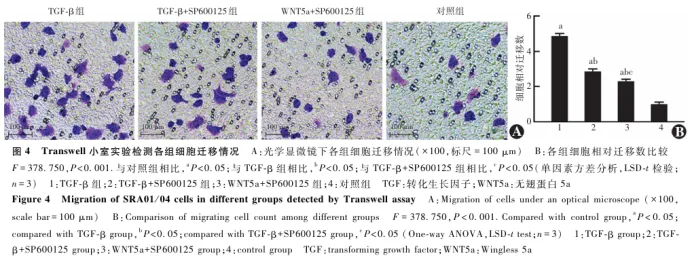
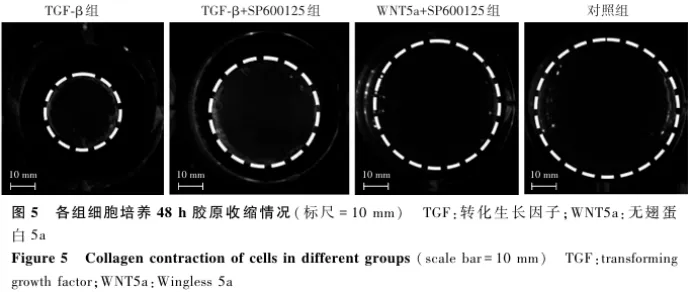
2.5 各组细胞Col-Ⅰ凝胶面积变化情况比较
Col-Ⅰ凝胶收缩实验结果显示,随着培养时间的延长,各组Col-Ⅰ凝胶面积逐渐缩小,培养48 h时,TGF-β组、TGF-β+SP600125组和WNT5a+SP600125组细胞Col-Ⅰ凝胶面积明显小于对照组,TGF-β+SP600125组和WNT5a+SP600125组细胞Col-Ⅰ凝胶面积明显大于TGF-β组( 图5 )。各时间点4个组细胞Col-Ⅰ凝胶面积比率总体比较差异有统计学意义( F分组=71.604, P<0.001),其中各培养时间点TGF-β组细胞Col-Ⅰ凝胶面积比率低于对照组、TGF-β+SP600125组和WNT5a+SP600125组,TGF-β+SP600125组低于对照组和WNT5a+SP600125组,WNT5a+SP600125组低于对照组,差异均有统计学意义(均 P<0.05);不同时间点各组细胞Col-Ⅰ凝胶面积比率总体比较差异有统计学意义( F时间=71.599, P<0.001)( 表3 )。
PCO是白内障术后患者视力下降的主要原因之一,可导致对比敏感度降低、眩光和复视,严重影响患者生活质量。PCO分为纤维性PCO和再生性PCO,前者与细胞过度增殖、细胞外基质(extracellular matrix,ECM)沉积收缩、细胞EMT有关,遵循经典的纤维化过程,后者主要表现为晶状体囊袋周围Soemmerring环和Elschnig珍珠,与晶状体纤维细胞的分化有关[3]。多种分子信号,如TGF-β信号通路、WNT信号通路等参与PCO的形成[6]。应激、炎症因子、G蛋白受体激动剂和促生长因子等通过胶原三螺旋重复序列包含蛋白1增强细胞膜上Rho家族小GTP酶活性,诱导膜近端蛋白丝裂原活化蛋白激酶激酶激酶(mitogen-activated protein kinase kinase kinase,MAP3Ks)的激活,磷酸化丝裂原活化蛋白激酶激酶4(mitogen-activated protein kinase kinase 4,MKK4)和MKK7,MKK7进一步磷酸化并激活JNK。活化的JNK激活转录因子激活蛋白1等下游分子,调控细胞增殖、分化、迁移、细胞自噬和凋亡,参与细胞纤维化[7,8]。WNT5a与FZD受体结合激活JNK,促进TGF-β诱导的ECM沉积。抑制WNT/JNK可减弱TGF-β对WNT5a的激活,减少细胞EMT和ECM沉积,抑制纤维化[9,10,11]。
在纤维化疾病中,TGF-β信号和非经典WNT信号激活相同的丝裂原活化蛋白(mitogen-activated protein,MAPK)通路,提示JNK与TGF-β、WNT信号紧密相连,共同促进组织器官纤维化[12]。细胞EMT和ECM沉积是纤维化的重要病理过程。Ras/TGF-β可促进细胞WNT5a/PCP(WNT/JNK)激活,诱导细胞EMT发生[13]。WNT5a/JNK 1通过磷酸化Paxillin、促进基质金属蛋白酶1(matrix metalloproteinases 1,MMP1)、MMP2、细胞黏附因子1和CD44的表达,上调波形蛋白和α-SMA的表达,降低E-钙黏蛋白水平,诱导胰腺癌细胞EMT,促进细胞迁移[9]。非经典WNT蛋白与FZD受体结合上调MMP7的表达,促进角膜上皮细胞迁移[14]。在敲除了Ⅱ型TGF-β受体(TBR-Ⅱ)的Tbr2 ceΔ/ceΔ小鼠中,TGF-β信号通路的异常可抑制JNK信号通路对角膜上皮细胞迁移力的促进作用,抑制角膜上皮的损伤修复[15]。PCO发生时,LECs转分化为肌成纤维细胞获得强迁移力,由晶状体囊袋赤道部游移至后囊下,形成瘢痕遮挡光学区[16,17]。α-SMA是肌成纤维细胞的标志物之一,参与细胞骨架的重塑,促进细胞迁移和收缩[18]。本研究发现,TGF-β刺激LECs中α-SMA蛋白表达显著增加,细胞转分化为肌成纤维细胞,抑制JNK磷酸化可有效下调细胞内α-SMA蛋白表达水平,阻断细胞EMT,抑制细胞迁移。
成纤维细胞、肌成纤维细胞分泌Col-Ⅰ、Col-Ⅲ和FN等ECM主要成分[19]。在细胞纤维化中存在大量不同类型的胶原蛋白,其中Col-Ⅰ为主要成分[19]。WNT蛋白可诱导LECs中Col-Ⅰ的表达,同时促进细胞收缩及瘢痕形成[20]。WNT5a与LECs的迁移、分化和ECM重构密切相关[21]。WNT5a与FZD8受体结合,激活JNK并诱导T细胞核因子活化和核转位,促进TGF-β诱导的ECM沉积。抑制WNT/JNK可减弱TGF-β诱导的Col和FN表达,逆转纤维化[22]。本研究表明,TGF-β促进LECs中Col-Ⅰ和FN合成,促进ECM沉积,抑制JNK磷酸化可有效抑制细胞Col-Ⅰ、FN的表达,减少ECM沉积,阻断LECs纤维化发生。
本课题组前期实验表明,沉默WNT5a表达后,TGF-β诱导的LECs纤维化得到显著抑制。WNT5a可作为TGF-β 1的下游作用位点,刺激肌成纤维细胞内Col、FN、波形蛋白表达,刺激ECM形成,诱导组织纤维化,沉默WNT5a可抑制TGF-β诱导的ECM沉积[10]。本研究结果也表明,TGF-β组细胞内Col-Ⅰ、FN和α-SMA表达显著增加的同时伴WNT5a、JNK、p-JNK的蛋白水平上调,经JNK抑制剂处理的细胞内Col-Ⅰ、FN和α-SMA表达显著降低,提示WNT/JNK信号通路参与TGF-β诱导的LECs EMT和ECM沉积。然而,本研究也存在一定的局限性,即仅分析了单一SRA0104细胞系在TGF-β/WNT5a/P-JNK信号轴诱导下的细胞变化。JNK信号通路在其他LECs细胞系和动物晶状体中对纤维化影响的确切机制有望成为下一步研究方向。
综上所述,本实验探索了TGF-β/WNT5a/JNK信号轴对LECs EMT和纤维化的促进作用,WNT5a/JNK信号通路可作为TGF-β信号通路下游作用靶点诱导LECs EMT,提高细胞迁移力,促进细胞Col-Ⅰ、FN、α-SMA的表达,促进ECM沉积,提高细胞收缩力,进而促进PCO的形成。JNK信号通路可能为PCO的药物防治靶点提供新思路。
利益冲突 所有作者均声明不存在利益冲突
作者贡献声明 宋宇:实施研究、采集数据、分析/解释数据、统计分析、起草文章;包秀丽:酝酿和设计实验、实施研究、指导研究、对文章的知识性内容作批评性审阅及定稿